ACTUAL ASPECTS OF THE PHILOGENESIS
АКТУАЛЬНЫЕАСПЕКТЫФИЛОГЕНЕЗА

Litvin V. V, Russia, PhD, Krasnodar, NANEB, KRC UNESCO
Литвин В.В., Россия, PhD, Краснодар, МАНЭБ, ККЦ ЮНЕСКО
Ключевые слова: Демография, продолжительность жизни, правило Орбели, элвилогия, лимит Хейфлика, правило М. Рубнера, теломер, метаболизм, микробиота, гипербарическая оксигенизация, плодовитость, долголетие.
Keywords: the demography, life expectancy, a rule of Orbeli, elvilogie, limit of Haflik, a rule of M.Rubner, telomer, a metabolism, microbiote, hyperbaric oxigenation, fruitfulness, longevity.
Аннотация:Человек вышел из животного мира (Ч. Дарвин) с горящей веткой в руках. Весь прошедший путь ознаменован стремлением жить долго, что стало реализацией инстинкта самосохранения на базе становления сознания. В результатенаблюдаетсяв основном монотонное повышение продолжительности жизни. При этом мировые полигинии демонстрируют активный демографический прогресс. Другие гендерные режимы стабильны или регрессируют. Однако в целом имеется рост, приближенный к геометрической прогрессии, что уводит Номо из естественного процесса развития биологического видового разнообразия на Земле. Согласно фундаментальному правилу Орбели СПЖ и плодовитость находятся для любого стабильного таксона в обратной зависимости. Снижение антропогенной нагрузки, определяемой объективными показателями Реймерса без ущерба для прогресса человечества сопряжено с целесообразностью смещения модуля распределения возрастной структуры социумов в сторону увеличения, что диктуется также задачами снижения преступности и более полного использования потенциала личности.
Археологические и антропологические данные, нарративы хронистов позволяют получить терминальную динамику продолжительности жизни человека от самых ранних стадий развития до современности.
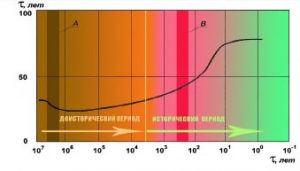
Рис.1. Динамика изменения средней продолжительности жизни рода НОМО.
Падение СПЖ в период 7-5 млн (зона А) лет назад связано с усилением и учащением военной деятельности древнего человека в связи с развитием сознания и морали.. Зона В – период средневековой европейской алхимии, в период существования которой в средние века было открыты свойства многих веществ полезных для лечения людей. Более того, с этого момента тема долгожительства уже не сходила с повестки дня работы в области медицины, психологии, биологических наук. Как обнаруживают на настоящий момент результаты исследований роль медицины, коммунального комфорта, кондитерского мастерства и противостоящей ему диетологии и нутриетологии и других достижений цивилизации практически исчерпана и деятельный процесс перешел в сферу психологии, генетики и биофизики. Успехи в этих науках определили ускоренный рост народонаселения Земли, что негативно сказывается на состоянии окружающей среды в глобальном масштабе.
Снижение антропогенной нагрузки на окружающую среду путем контроля демографического прогресса может существенно деформировать социальную среду сообществ, связанную с торможением креативных функций социума и катастрофическому процессу деградации производительных сил, некомпенсируемой развитием искусственного интеллекта. Тогда с учетом правила Орбелли необходимо переходить к контролю структуры самого социума. Известно, что жизненные силы видов покоятся на фундаменте потоков и сродства, что в соответствии с теорией взаимности Онзагера с очевидной необходимостью приводить в соответствие желаемым потокам соответствующие им сродства. С точки зрения производства энтропии популяцией правило Орбели таким образом представляет собой частный случай теории взаимности Онзагера.

Рис.2. Прогностические оценки численности населения с учетом числа детей в семье (С.Щербаков).
Однако кроме количественных оценок должны быть приняты во внимание и качествен-ные. Кроме того, спектр биологических оценок членов сообществ, охватывающих важнейшие социально-значимые характеристики и может стать основой для программной деятельности в перспективе.
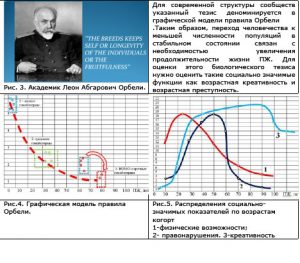
Рис. 3. Академик Леон Абгарович Орбели. Для современной структуры сообществ указанный тезис деноминируется в графической модели правила Орбели
.Таким образом, переход человечества к меньшей численности популяций в стабильном состоянии связан с необходимостью увеличения продолжительности жизни ПЖ. Для оценки этого биологического тезиса нужно оценить такие социально значимые функции как возрастная креативность и возрастная преступность.
Рис.4. Графическая модель правила Орбели.
Рис.5. Распределения социально-значимых показателей по возрастам когорт
1-физические возможности;
2- правонарушения. 3-креативность
Совершенно очевидно, что для человечества предстоит в будущем иметь структуру социумов с преобладанием индивидов в возрасте более 50 лет. Члены такого социума более законопослушны и обладают максимальным креативным потенциалом.В силу этого перед наукойстоит задача исследовать пути достижения деятельного и здорового долголетия. При этом должны быть использованы методы Элвилогии.
Общие закономерности процессов, происходящих в больших популяциях, обладающих относительной стабильностью в течение значительного периода времени с термодинамической точки зрения контролируются не только внутренним явлениями, присущими ансамблю, но и видовыми свойствами, определяющими энергообмен с внешней средой, что характеризует открытую систему. Вероятность состояний, в которых может находиться популяция, может быть определена как вероятность независимых совместных явлений в течение эквивалентного времени по всем допустимым диапазонам биологических процессов статистического ансамбля:
W = W1W2 W3 …Wn
где:
W- вероятность состояния популяции,
1.2,3,… n – индексы сопряженных одновременных проявлений системы, характеризующие ее внутренние свойства.
Для термодинамической оценки популяции как системы с таких позиций справедливо простое неравенство, названное нами границей большого круга:
n m r
S(t) = k ln П W i + k ln П W j + S g(t) +
i=1 j=1 g=1
g z= N(t)
+ S v (t) — Sqz(t) dt 0
v=1 z=1t
где:
S(t) — энтропия, производимая системой за эквивалентное время, Дж/ К,
i – индекс произведения вероятностей всех возможных проявлений в популяциях -появлений индивидов в ансамбле, их взаимодействий и жизнедеятельности.
j – индекс произведения вероятностей потери жизнеспособности индивидами,
g – индекс процессов деятельности,
v – индекс метаболических процессов роста,
q – индекс энергообмена
t- текущее время
z- индекс суммирования по числу индивидов популяции
k- коэффициент энергетической эквивалентности, постоянная Больцмана,
При детальном рассмотрении v-процессов в отношении их аддитивных энтропийных вкладов, то в общей сумме они оказываются незначительными. Аналогичная оценка может быть дана и для g-процессов.
Модель для статистического ансамбля с числом доступных состояний W, в случае с биологическими объектами, имеет постоянную k, характеризующую и вероятностные и энергетические свойства системы, поэтому опирается на атомистический уровень модели Больцмана, хотя «частицы» рассматриваемых статистических совокупностей здесь вступают во взаимоотношения, характеризующие их специфические свойства, отличные от газовых.
Представленная модель может быть корректной для популяций с достаточно большим числом индивидов или особей так как она имеет аналогичное статистическое содержание, которое имел ввиду в 1877 году Людвиг Больцман (1844-1906). Для такого термодинамического анализа нас будут интересовать гомойотермные виды, для которых характерна стабильная температура тела, что не просто облегчает процесс вычислений, а и позволяет рельефно выявить видовые различия с энергетической точки зрения. Гомологическая близость этих видов к человеку дает нам надежду на более основательную оценку полученных результатов, применительно к главной теме. Некоторые закономерности энергообмена видов известна. Например, у видов с малой видовой продолжительностью жизни теплопроизводство особей выше, чем у видов с большей ВПЖ, что является содержанием правила Макса Рубнера (1894).
Для расчета приняты некоторые вполне реальные допущения, не искажающие эволюционных свойств видов. Популяции численно стабильны, то есть число потерявших жизнеспособность равно числу родившихся, то есть имеет место простое воспроизводство. Для удержания популяций в заданной численности видам приходится производить разное число детенышей, при этом соотношение между численностью популяции N и когортой пополнения No за эквивалентное время to составит величину:
Тmax
N/ No = F(t) d t = const,
о
где: F(t) – функция распределения численности особей по возрастам от 0 до tmax.
С некоторой долей приближения можно использовать функцию Б. Гомперца (1825) и В.Мэйкхэма (1860), при этом может возникать некоторая погрешность, так как это приближение плохо работает на периферии распределения. Для популяций людей при возрастах до 20 лет получаются завышенные, а при возрастах более 70 лет — заниженные оценки численностей когорт.
t
No = N / { exp R/[1-exp (t) ]+ А t }d t
0
где:
А – невозрастная (0,01, 1/год), R и — возрастные константы уравнения математической модели потери жизнеспособности (0,00005 1/год и 0,1 1/год соответственно).
Второе допущение, принятое в построении термодинамической модели популяций состоит в том, что число женщин и число мужчин в диапазоне репродуктивного возраста равны, аналогичное равенство имеет место и для других видов. Распределение числа родившихся по возрастам рожениц взяты из демографических справочников ООН.
В качестве исходных данных для расчетов приняты средние значения показателей, характеризующих базовые виды: человек, собака и крыса. Такой выбор видов обусловлен близкими условиями жизнедеятельности и контрастными свойствами в смысле различного распределения вероятностей потери жизнеспособности по возрастам, что отражается на величинах значений интегралов распределения Гомперца — Мэйкхэма.. Величина интеграла принята в пределах, характерных для стабильных популяций европейских стран.
Здесь следует отметить, что для многих популяций животных и птиц, живущих в естественных условиях, интеграл Гомперца-Мэйкхэма имеет значения существенно меньшие, чем приведенные. Это связано с давлением среды, и прежде всего с влиянием хищников, резко снижающих численность молодняка. Если устранить эти влияния, то есть сохранить собственно способность вида к реализации, заложенной в нем долговечности, то отличия видов будут не так разительны, как это имеет место в сложившихся ареалах. Для себя человек создал условия минимального влияния других видов, но имеет другие внутривидовые ограничения.
Другими словами люди начинают интенсивно терять жизнеспособность с окончанием детородного периода (55 – 60 лет) Аналогичные распределения имеют место и для других видов. У собак, крыс, многих животных сохраняется подобная закономерность. Такова логика эволюции — давший жизнь потомству, уже выполнил свой долг перед видом.
Итак, зная распределение чисел особей по возрастным группам, можно определить величину видового показателя, характеризующего изменение энтропии потери жизнеспособности.
Реализация актов поддержания жизнеспособности отдельных особей, принадлежащих всей совокупности как независимых событий, может осуществляться большим числом вариантов i. Термодинамическая вероятность этих процессов определится как функция численности равновесной популяции. Энтропию системы, возникающую вследствие рождений, можно оценить путем расчета числа доступных состояний системы, исходя из вероятности появления детей (детенышей) в соответствующих возрастных группах женщин (самок) репродуктивного возраста, которую можно определить по таблицам рождаемости. Несмотря на то, что появление детей является в большинстве случаев планируемыми событиями, они в конечном итоге взаимно независимы и носят случайный характер. Определяя так вероятность появления детей (детенышей), можно рассчитать число способов реализации состояния системы в процессе воспроизводства. Расчеты показывают, что число доступных состояний последнего намного меньше общего числа доступных состояний жизнедеятельности всей популяции, что оценивается произведением i. В самом деле, число особей женского пола в репродуктивном возрасте намного меньше численности членов всей популяции, совершающих движения в ансамбле.
В предельном случае основное неравенство представляется уравнением баланса приращения энтропии, произведенного за эквивалентное время всей системой, из которого можно получить значение показателей Lv (линия жизни) для каждого отдельно биологического вида.
n=N
klnN! – kzln ( N/z)! = Sqn(t) dt ,
tn=1
100
где: z = Lvo/Lv, a Lvo = { exp R/[ 1-exp (t) ] + А t }d t
0
Из этого уравнения следует, что видовая характеристика Lv определится из зависимости типа уравнения Аррениуса в энтропийных терминах:
Lv = Lvo exp (- S/k),
где: S – энтропия активации, Дж/К
Известно, что удельное массовое производство энтропии в организмах не является постоянной величиной в течение развития, роста и старения. Эта величина в ходе онтогенеза непрерывно падает, что исследовано многочисленными авторами и обобщено А.И. Зотиным. Непостоянна она и течение суток и варьируется по особям. Оценку интегральной суммы
n=N
Sqn(t) dt ,
tn=1
можно осуществить по среднему уровню обменной энергии. При этом расчетные значения S соотнесены с размерностью константы Больцмана (Дж/К), с учетом количества постоянных молекул в клетке (протеинов, нуклеиновых кислот, фосфолипидов и других высокомолекулярных соединений), в среднем – это 100 млн и их молекулярной массы – 1 млн. Количество механической энергии, затрачиваемой скелетными мышцами на производство работы во внешней среде, незначительно по сравнению с общей теплопродукцией, что отражено в расчете.
Самое интересное в этой общей картине – сравнение видовых величин Lv, и определение общей тенденции их связи с продолжительностью жизни особей соответствующих видов.
Таблица показателей видов животных и человека и расчетное значение видовой характеристики Lv.
№ вид продолжи-тельность жизни, лет Теплопродук-ция, Вт/кг Эквива-лентное время, сутки Lv lg Lv
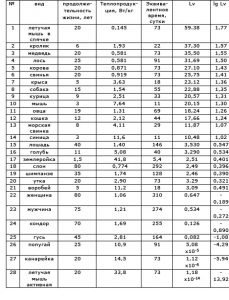
Если развернуть результаты расчета в координатах: удельная энергия обмена – продолжительность жизни видов, то линии равных значений Lv образуют семейство гипербол, среди которых находится гипербола правила М. Рубнера:
qt = 1,75 x 106 , кДж/кг,
являющаяся частным случаем общего семейства:
qt = kT(lnLvo – lnLv)х100, кДж/кг.
Сам Рубнер определял значение qt = 8,37х105 кДж/кг, что впоследствии было уточнено.
Здесь, для биологических систем постоянная Больцмана как и в случае неживой природы стала универсальным мостом между свойствами частиц системы и свойствами всей системы.
Вот практический пример расчета возможности увеличить продолжительность жизни человека. При предполагаемой продолжительности жизни мужчин, оцениваемой в 75 лет:
Lv = Lvoexp (- S/ K) = 69,06 х ехр(-4,86) = 0,534
Если удастся снизить интенсивность метаболизма на 15%, с 1,209 Вт/кг до 1,028 Вт/кг, что представляется вполне осуществимым, то из генеральной совокупности
q t = kT(ln Lvo – ln Lv)100,
тогда:
t = kT(lnLvo – lnLv)100/q = 1,38х10-23х1,42х1029х309 (ln 69,06 – ln 0,534)/1,028 = = 2,860 х109с = 90,7 года.
Известная тенденция продления жизни посредством замедления хода биологических часов, через снижение скорости метаболических процессов в данном случае оценена с позиций термодинамической вероятности. Прибавка на более, чем 15 лет не помешала бы многим желающим.
Однако кроме снижения энергетических показателей в последнее время наука получила и передала в практику новые технологии увеличения продолжительности жизни. Это относится к методу «трансплантации» микробиоты. Идеология способа исходит из того, что в ЖКТ любого человека спонтанно организуется ансамбль сапрофитов, при этом не всегда благоприятный. Долгожительство в некоторой степени связывается с деятельность этого ансамбля. Опыты пересадки ансамблей от долгожителя к реципиенту дают положительный эффект.
Известно, что каждый акт пролиферации сопряжен с укорочением конечных участков ДНК, теломеров, что определяет лимит Хейфлика. Применение ферментов для восстановления теломеров — теломеразы не дали стабильного результата. Ученым Израиля удалось найти способ достройки теломеров с помощью гипербарической оксигенации.
ЛИТЕРАТУРА
1. Литвин В.В. Мы или как прожить 150 лет, Ростов Дон, Феникс:2004, -537 с.
2. Литвин В.В. Термодинамический аспект в расчетах долговечности/8-й Congress on Materia lTesting, OMIKK-Technoinform, Budapest,1982, Lectures, 1.Volume,234-237pp
3. Литвин В.В. ЮНЕСКО на кубанской земле,/Журнал Вестник ККЦ ЮНЕСКО, 2000, вып.1, с 3-8.
4. Ноздрачев А.Д. и др. Начала физиологии, СПб: Лань, 2001, -1088 с.
5. Фролькис В.В., Мурадян Х.К. Экспериментальные пути продления жизни. Л.: Наука 1988, -248 с.
6. Дильман В.М. Большие биологические часы, М: Знание, 1981. -208 с.
7. Пицхелаури Г.З. Лет до ста расти. М: Знание,1982,- 96 с.
8. Курцмен Дж., Гордон Ф. Да сгинет смерть, М: Мир, 1987 -222 с.


